

Endowiki: aggiornamento continuo di endocrinologia
Per tutti gli operatori di endocrinologia: lo stato dell'arte su clinica e terapia.
Uno strumento semplice e fruibile da tutti coloro che intendono essere sempre aggiornati, promosso dai migliori operatori del settore.
News
Inquadramento generale diagnostico delle patologie gonadiche femminili

Anatomia dell'ovaio
Maurizio Poggi, Chiara Pascucci
UOC di Endocrinologia, Azienda Ospedaliera Sant’Andrea, Roma
Le ovaie o gonadi femminili, sono ghiandole pari, ubicate vicino alla parete laterale del piccolo bacino. L’ovaio ha una posizione variabile, in quanto può seguire gli spostamenti dell’utero, sia in corso di gravidanza che a seguito di gravidanze pregresse. Nelle multipare l’ovaio assume spesso una posizione più bassa e posteriore, limitata davanti dall’uretere, indietro dalla parete pelvica posteriore e in basso e medialmente dal legamento utero-sacrale. L’ovaio non è ricoperto dal peritoneo viscerale, ma da un epitelio particolare, detto epitelio ovarico o germinativo (in passato si riteneva che vi originassero i gameti durante l’ontogenesi, ma in realtà le cellule germinali hanno origine fuori dalla gonade, nell’entoderma del sacco vitellino, e migrano in essa durante lo sviluppo), che origina dal rivestimento della cavità celomatica primitiva. Le ovaie hanno la forma di un ovoide appiattito, con il maggior asse verticale disposto su un piano pressoché sagittale, e consistenza dura con una superficie irregolare.
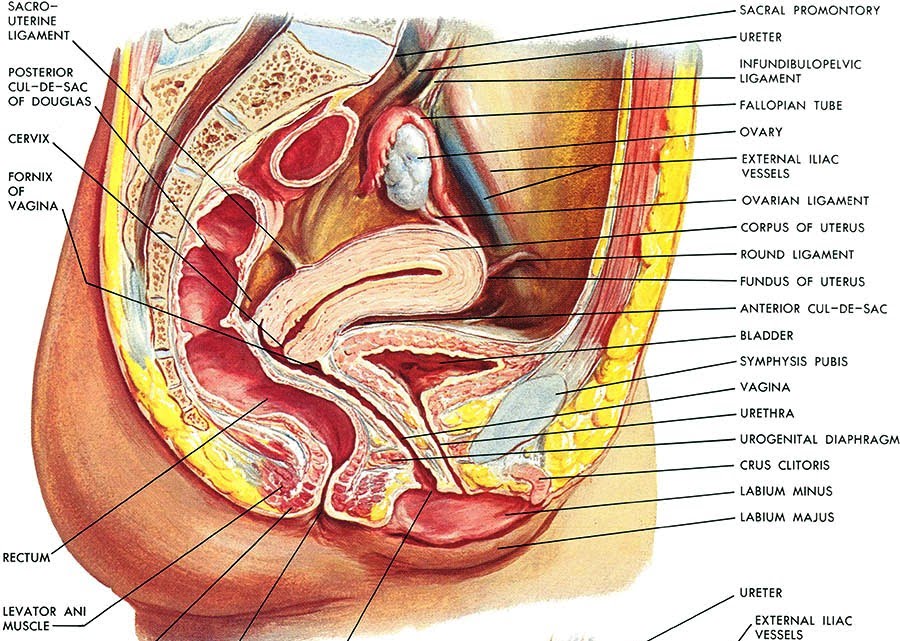
Durante il periodo fertile le dimensioni dell’ovaio sono: altezza 3.5 cm, larghezza 2 cm, spessore 1 cm. L’ovaio delle pluripare è più grosso di quello delle nullipare. Sia nelle bambine che durante la vecchiaia l’organo risulta più piccolo. Nel primo caso non è ancora avvenuto lo sviluppo, mentre in vecchiaia si verificano fenomeni di sclerosi.
L’aspetto dell’ovaio è variabile a seconda delle fasi della vita sessuale: prima della pubertà la superficie si presenta liscia e regolare; durante il periodo fertile assume un colorito grigio-rossastro e diventa notevolmente irregolare per la sporgenza di follicoli in via di maturazione e di corpi lutei. Con il progredire dell’età compaiono numerose cicatrici, esito dei processi di atresia follicolare e della regressione dei corpi lutei, mentre in menopausa l’ovaio tende a diventare atrofico, acquistando una consistenza quasi lignea. Infine, dopo i 70 anni la superficie dell’ovaio ritorna ad essere liscia per atrofia delle cicatrici stesse.
L’ovaio è mantenuto in sede da mezzi di fissità e precisamente da 4 legamenti: il legamento sospensore o lombo-ovarico, l’utero-ovarico, il mesovario e il legamento tubo-ovarico.
L’ovaio ha una duplice fonte di irrorazione: l’arteria genitale (ovarica) e il ramo ovarico dell’arteria uterina. L’arteria genitale nasce dall’aorta addominale, a livello della seconda vertebra lombare, a destra passa davanti alla vena cava inferiore e incrocia da ambo i lati l’uretere e il tratto iniziale dei vasi iliaci esterni, raggiunge l’estremità superiore dell’ovaio penetrando nel mesovario e dando origine ai rami che penetrano nell’ilo e si distribuiscono al parenchima ovarico. Il ramo ovarico nasce dall’arteria uterina e seguendo il legamento utero-ovarico raggiunge il mesovario per anastomizzarsi con l’arteria ovarica. Le vene formano un ricco plesso in corrispondenza della zona midollare dell’ovaio, vanno a formare il bulbo dell’ovaio e fanno capo alla vena uterina e alle vene ovariche. Le vene ovariche sono riunite in un plesso pampiniforme che risale verso l’addome nel legamento sospensore, poi convergono in un unico tronco che sbocca a sinistra nella vena cava inferiore e a destra nella vena renale. I linfatici efferenti dell’ovaio fanno capo ai linfonodi pre-aortici e para-aortici.
I nervi formano un ricco plesso attorno alle arterie. La zona della midollare dell’ovaio è ricca di fibre nervose, che seguendo il decorso dei vasi giungono alla zona corticale. Il plesso utero-ovarico proveniente dal plesso celiaco accompagna l’arteria genitale e fibre nervose provenienti dal plesso pelvico seguono il ramo ovarico dell’arteria uterina.
La zona corticale dell’ovaio è la parte più periferica, situata sotto l’epitelio ovarico. Ha uno spessore compreso dai 2 ai 10 mm ed è formata dallo stroma corticale, tessuto connettivo particolare dotato di un notevole polimorfismo e di una notevole capacità d’adattamento funzionale. Lo stroma contiene follicoli oofori e corpi lutei in vari stadi d’evoluzione o di regressione e cellule in grado di produrre ormoni androgeni. Sotto l’epitelio ovarico che ricopre la corticale, vi è un ricco strato di fibre collagene con scarse cellule che formano la falsa albuginea. La zona midollare è la zona più profonda e centrale dell’ovaio, in continuazione dell’ilo attraverso il quale decorrono i vasi sanguigni, linfatici e i nervi che si distribuiscono all’ovaio. La midollare è costituita da tessuto connettivo lasso che contiene fibre elastiche e fasci di fibre muscolari lisce che accompagnano le ramificazioni delle arterie. Queste ramificazioni vascolari presentano una disposizione a spirale caratteristica, che conferisce alla zona profonda della midollare una certa capacità di deformarsi in rapporto allo stadio di sviluppo dei follicoli situati nella corticale e a modificare la sua consistenza in rapporto allo stato di congestione delle arterie. Nella midollare troviamo anche vasi linfatici e corpi albicanti vascolari, che sono formazioni ialine risultato del processo di degenerazione della parete dei vasi ovarici. Nella zona dell’ilo troviamo cellule dalla forma allungata, fornite di un grosso nucleo e di un citoplasma acidofilo capaci di produrre androgeni.
Le ovaie svolgono il duplice compito sia di produrre oociti (funzione gametogenica) sia di secernere estrogeni, progesterone e una percentuale degli androgeni circolanti (funzione endocrina). I compartimenti in grado di sintetizzare steroidi sono:
- follicolo
- corpo luteo
- cellule interstiziali
Il follicolo è l’unità morfo-funzionale dell’ovaio. Può trovarsi in fase quiescente (follicoli primordiali) o in fase di maturazione. Già al momento della nascita la femmina possiede il suo completo patrimonio follicolare, che consta di circa 1 milione di follicoli primordiali; questo patrimonio va poi riducendosi, passando dalla nascita alla pubertà. Dal menarca alla menopausa solo 400-500 follicoli andranno incontro a completa maturazione e all’ovulazione, mentre tutti gli altri follicoli andranno incontro ad involuzione atresica.
I follicoli primordiali sono costituiti da un oocita circondato da uno strato di cellule appiattite rivestite da una lamina basale. L’oocita, del diametro di 25-30 µm, si trova bloccato nella profase della prima divisione meiotica. Le membrane plasmatiche dell’oocita e delle cellule follicolari sono strettamente adese fra loro, grazie a sistemi di giunzione detti desmosomi. Questi follicoli primordiali sono in fase quiescente e possono sia evolvere allo stadio primario che andare verso la regressione. La lamina basale separa le cellule follicolari dallo stroma circostante.
I follicoli secondari presentano cellule follicolari disposte in più strati attorno all’oocita, che continua a crescere. Tra le cellule follicolari compaiono piccoli spazi dove si accumula un po’ di liquido. La membrana plasmatica dell’oocita inizia a presentare delle villosità, che si ingranano con i prolungamenti citoplasmatici provenienti dalle cellule follicolari che lo circondano. Tra l’oocita e le cellule follicolari si viene a formare la zona pellucida. Si ha un’organizzazione del connettivo peri-follicolare in strati concentrici. I follicoli secondari migrano nella zona profonda, mentre i follicoli primordiali nella superficie della zona corticale.
Nei follicoli cavitari gli spazi tra le cellule follicolari confluiscono, formando una cavità unica che contiene il liquido follicolare secreto dalle cellule follicolari. L’accrescimento progressivo di questa cavità spinge le cellule follicolari alla periferia dove esse formano la parete del follicolo, costituita da pochi strati di cellule che per le loro caratteristiche sono state chiamate cellule della granulosa. Attorno all’oocita, che ha raggiunto il diametro definitivo di 125-150 µm, la granulosa forma una sporgenza chiamata cumulo ooforo. L’oocita è circondato dalla zona pellucida dello spessore di circa 5-10 µm. Le cellule che circondano l’oocita con interposizione della zona pellucida, hanno disposizione raggiata e formano la corona radiata. Intorno all’oocita si formano le guaine tecali. Le cellule dello stroma peri-follicolare si modificano, la sostanza inter-cellulare tra di esse si riduce notevolmente e rimane rappresentata soltanto da una trama di fibrille reticolari che accompagna una ricca rete capillare. La teca interna così formata ha cellule in grado di produrre androgeni. La teca esterna assume un aspetto più fibroso e vi si riscontrano alcune fibrocellule muscolari lisce.
I follicoli maturi hanno un diametro di 12-15 mm. Normalmente si ha un solo follicolo maturo per ciascun ciclo mestruale. La parete follicolare è sottile e il follicolo ripieno di liquido ha un aspetto cistico e sporge dalla superficie dell’ovaio. La porzione del contorno follicolare che sporge è molto sottile e trasparente ed è denominata stigma. A livello dello stigma avviene la rottura del follicolo e l’oocita circondato dalle cellule del cumulo ooforo viene espulso (follicolo deiscente). Alla deiscenza follicolare concorrono diversi meccanismi: la produzione di enzimi litici da parte delle cellule dell’epitelio superficiale e della teca, l’edema della parete follicolare, la contrazione di elementi muscolari peri-follicolari e la congestione dei vasi midollari. Nei follicoli maturi l’oocita porta a termine il suo accrescimento e completa la prima divisione meiotica, dando origine al primo globulo polare e all’oocita di secondo ordine, nel quale il corredo cromosomico è ormai aploide. Lo sblocco della meiosi è dovuto alla neutralizzazione da parte dell’LH di un fattore inibente la meiosi prodotto dalle cellule del cumolo ooforo e reperibile nel liquido follicolare. Al momento della sua espulsione l’oocita è bloccato nella metafase della seconda divisione meiotica. La progressione del follicolo attraverso i vari stadi di maturazione è dettata dall’azione del FSH. La produzione degli estrogeni da parte della teca interna, lo scoppio del follicolo e la successiva trasformazione delle sue pareti in corpo luteo sono invece dovuti all’azione dell’LH.
I follicoli involutivi sono tutti quelli che non raggiungono la completa maturazione e si arrestano a un stadio qualunque del loro sviluppo, con conseguente regressione e atresia follicolare. L’atresia follicolare deriva dalla necrosi dell’ovocita e delle cellule della granulosa, con ipertrofia delle cellule della teca che si trasformano in cellule interstiziali secondarie.
Le cellule interstiziali si distinguono a seconda dell’origine in cellule tecali-interstiziali e in cellule interstiziali secondarie.
Le cellule tecali-intersiziali sono localizzate nella teca interna dei follicoli e secernono androgeni, soprattutto androstenedione. Queste cellule acquisiscono attività enzimatica 3ßidrossi-steroido-deidrogenasi delta-isomerasi e di clivaggio della catena laterale del colesterolo, nonché i recettori per l’LH. Sotto lo stimolo dell’LH che giunge attraverso i capillari tecali, si ha l’evoluzione di queste cellule interstiziali in grosse cellule epiteliali capaci di secernere progesterone. Successivamente acquisiscono l’enzima 17alfa-idrossilasi C17-20 liasi e passano dall’essere cellule progestinico-secernenti a cellule androstenedione-secernenti.
Le cellule interstiziali secondarie si hanno allo stadio di atresia follicolare e si formano per l’ipertrofia delle cellule tecali interstiziali e continuano a produrre androgeni sotto lo stimolo dell’LH.
Il Corpo Luteo è la trasformazione, dopo l’ovulazione, del follicolo. La lamina basale, dopo la rottura del follicolo, si frammenta consentendo la penetrazione tra le cellule della granulosa dei fibroblasti e dei vasi della teca all’interno della cavità follicolare con la liberazione di citochine ed altri ligandi. Si forma così un coagulo che si organizza prendendo l’aspetto di un essudato connettivo circondato da un ammasso di emazie, fibrina e macrofagi ripieni di emosiderina. La penetrazione tra le cellule della granulosa dei vasi provenienti dalla teca interna è responsabile della trasformazione delle cellule della granulosa in cellule luteiniche di origine granulosa che producono progesterone. Nello stadio di maturità il corpo luteo appare come una formazione giallastra sporgente sulla superficie dell’ovaio. Le cellule della teca luteiniche esprimono invece l’enzima P450 17 a idrossilasi e dopo la luteinizzazione producono androstenedione che, grazie alla capacità aromatasica del corpo luteo viene trasformato in estradiolo. In caso di mancata fecondazione il corpo luteo va incontro a un processo di luteinizzazione che inizia otto giorni dopo l’ovulazione ed è secondario alla regressione delle cellule della teca, residuando così un ammasso di tessuto connettivo definito corpus albicans.
Bibliografia
- Monaco F. Endocrinologia Clinica. Società Editrice Universo, V Edizione.
- Balboni GC, et al. Anatomia Umana. Edi-Ermes, Volume II, III Edizione.
Login Form
Area Pubblica

Main sponsor

